

Le langage des dauphins
I. Un animal doué de langage ?
Un texte prophétique de R.Stenuit
Une question toujours controversée
Les très cruelles recherches du Dr John Lilly
Les recherches à Hardewijk
Les recherches en Mer Noire de Vladimir I. Markov
Cymascope : comprendre le langage dauphin
L’espéranto chez les dauphins
II. Voir et parler : une même fonction
Echogrammes et transmission d’images
Sons purs et signatures sifflées
Infrasons et autres productions sonores
III. Une linguistique universelle
Parler aux extraterrestres : le Projet SETI
IV. Références

Un monde sonore
I. Un animal doué de langage ?

Un cerveau complexe pour une cognition complexe…
A. Une question toujours controversée
L’attitude du corps scientifique reste encore extrêmement réservée quant à l’existence éventuelle d’un langage chez les cétacés. On s’en doute, le fait de reconnaître une compétence cognitive aussi pointue chez un animal non-humain s’avère douloureux pour nos certitudes anthropocentristes. L’ultime rempart entre l’animalité et l’humain se réfugie en effet dans cette capacité prétendument unique si l’on en croit Noam Chomsky et la plupart des linguistes- que nous aurions d’émettre des sons vocalisés articulés en chaînes, porteurs de sens, et s’organisant selon une grammaire propre.
Il est curieux que beaucoup d’humanistes actuels s’en tiennent farouchement à ce besoin d’une rupture ontologique nette entre l’animal et ce qui, selon eux, n’en est plus un : l’homme. Toutes sortes de critères ont servi à cette fin : le rire, la guerre ou la culture ont également été désignés en leur temps comme des sommets inaccessibles à la «bête brute».
Pour J.H. Fabre par exemple, (in « Souvenirs entomologiques ») la supériorité écrasante de l’être humain tient non pas tant à ce qu’il parle mais à ce qu’il est « le maître de l’outil et du feu ». Depuis lors, il est apparu que nombre d’espèces animales se servaient bien d’outils, et quant au feu, on voit mal quel usage pourraient en faire les grands mammifères marins.
Force est d’admettre, par ailleurs, que tout animal communique avec ses semblables. On sait que les bonobos font grand usage d’une gestuelle subtile, on sait que les éléphants se servent d’infrasons transmis par le sol et recueillis avec la trompe – pour échanger des informations à des distances énormes, on sait que singes verts, mangoustes, perroquets ou corbeaux font usage de « cris codés » pour désigner des choses ou des animaux (un aigle n’est pas annoncé de la même manière qu’un serpent), etc.
Mais à cela, bien sûr, les tenants du monopole humain répondront qu’il ne s’agit que d’échanges simples, portant sur des émotions brutes ou des désirs immédiats, trahissant la colère ou manifestant quelque stratégie sociale, mais qu’en aucun cas, il ne s’agit de « vrai langage » puisque cet outil ne peut servir à conserver la mémoire du passé, à désigner des choses absentes ni à évoquer le futur.
A vrai dire, aucune étude scientifique ne nous permet d’affirmer ou d’infirmer cette assertion. Pour ce qui est des éléphants, par exemple, nous savons qu’ils sont dotés de cultures diverses qui se transmettent par l’enseignement, mais nous ignorons encore de quelle manière précise ces informations sont transmises et si le «parler par infrasons» pratiqué par ces pachydermes comprend des mots, des vocables ou de la syntaxe.
En revanche, et c’est ce que nous allons étudier tout au long des pages qui suivent, une quantité sans cesse croissante de recherches menées tant en laboratoire que sur le terrain, nous amène aujourd’hui à une conclusion qui semble inexorable en termes scientifiques :
Oui, les grands cétacés disposent d’un système de vocalisation à ce point sophistiqué qu’il peut, sans doute aucun, être appelé un «langage articulé» au sens propre du terme.
Même s’il ne comporte sans doute aucun mot, mais bien d’autres éléments syntagmatiques…

Dauphins tachetés (Stenella frontalis) autour de Bimini aux Bahamas. Le Dr Denise Herzing étudie cette population depuis 30 ans. Une grande partie de son travail consiste à tenter de créer une communication bidirectionnelle avec les cétacés.
B. L’expérience de Bastian
Richard C. Connor, dans un ouvrage déjà ancien intitulé Lives of Whales and Dolphins, exprime l’opinion la plus courante sur cette question, celle que l’on retrouve aujourd’hui dans la plupart des guides cétologiques ou des ouvrages de vulgarisation consacré aux dauphins :
« Afin de découvrir l’existence d’une langue parlée chez les dauphins, explique Connor en substance, les scientifiques ont tenté d’interpréter les messages contenus dans leurs vocalisations. Sans doute ne pouvaient-ils en déchiffrer les codes mais il est évident qui si un tel langage existe, il doit nécessairement comprendre un nombre important de vocalisations variées et différentes. Cela ne semble pas être le cas. Les émissions sonores des dauphins sont actuellement envisagées comme de simples cris inconscients, tels qu’en produisent tous les mammifères, mais la question reste toujours pendante. Une chose est en effet apparue évidente lorsque nous avons enregistré les vocalisations des Tursiops de Shark Bay. C’est que les clicks et autres « sons pulsés » sont infiniment plus variés que nous ne le soupçonnions. Ce type d’émissions sonores représente donc à cet égard un riche terrain d’investigation ».
Lors de la fameuse expérience menée par Jarvis Bastian entre 1964 et 1968, continue Connor, on a voulu savoir si les dauphins étaient capables de se transmettre des informations de type conceptuel, telles que «à gauche » « à droite », « en haut » « en bas » « éteint » ou « allumé », et ceci sans apprentissage, à partir d’un langage supposé «naturel».
Récompensés par des morceaux de poissons – cette pratique est aujourd’hui contestée, car elle crée de la confusion dans l’esprit de l’animal – le dauphin Buzz et sa compagne Doris furent enfermés dans un même bassin. Un écran opaque tirés d’un bout à l’autre de la piscine empêchaient qu’ils se voient ou qu’ils n’entrent en contact.
Pour obtenir sa récompense, chaque dauphin devait sélectionner l’une de deux pédales d’expérimentation que l’on avait placé face à lui. La femelle était avertie du choix de la bonne pédale, mais pas le mâle, de sorte que si un langage existait, on pouvait à bon droit supposer que les cétacés en feraient usage pour s’informer mutuellement.
Le mâle n’était donc sensé presser la « bonne » pédale que lorsque la femelle l’avait fait avant lui.
Les seules informations disponibles pour lui étaient d’ordre acoustique, l’écran central empêchant tout contact visuel.
Au grand soulagement des chercheurs, oserait-on dire, les résultats furent décevants : les dauphins n’échangèrent que quelques « sons pulsés » élémentaires. Certains messages furent peut-être transmis d’un individu à l’autre par la seule voie des émissions sonores, mais il fut désormais considéré comme acquis – sur base, rappelons, d’un unique protocole d’expérience répétée par deux fois (Bastian, Wall and Anderson, 1968.) – que les dauphins ne disposaient d’aucune langue naturelle.
Sans doute fallait-il s’y attendre : Buzz et Doris avait été capturés à l’âge d’un an et n’étaient respectivement âgés que de 3 ans et 4 ans lors des premières expériences, conduites en outre dans un contexte technologique relativement primitif.
Richard Connor conclut en effet le récit de cette expérience par ces mots : « Il n’y a donc aucune preuve que les dauphins usent d’un langage entre eux » dit-il « mais il est vrai que d’énormes avancées ont été faites en ce domaine durant ces plus récentes années avec l’avènement des micros subaquatiques et des méthodes de « tracking ». Peut-être un jour, avec le temps, les scientifiques
seront-ils alors capables de découvrir d’autres indices à propos de ce que ces créatures se disent et plus encore, sur le contenu de leurs communications ».
Voir ci-dessous les résultats positifs de l’expérience de Bastian à Hardewijk.
Lire le récit complet de l’expérience en anglais sur le site de Ken Levasseur
Sommaire
C. Les recherches sur le langage au delphinarium de Harderwiik

Harderwijk dans les années 60
« Étant donné le très grand développement du cerveau des odontocètes, on s’est demandé dans quelle mesure ces animaux pouvaient communiquer entre eux. Dés que l’on a entrepris de conserver des dauphins souffleurs (Tursiops truncatus) en captivité au delphinarium de Marineland en Floride, le premier qui ait été construit au monde, ces animaux ont donné l’impression qu’ils conversaient entre eux.
En 1965, nous avons gardé, dans notre propre delphinarium, un dauphin souffleur isolé pendant deux mois.
Un tuyau de polyester de 10 centimètres de diamètre servait de déversoir entre son petit bassin et le grand bassin où séjournaient les autres animaux ; les deux extrémités en étaient immergées. Nous pouvions fréquemment voir un dauphin à chaque bout de ce tuyau: par des sons clairement audibles pour nous qui étions hors de l’eau, ils établissaient le contact un avec autre. Souvent, un animal du grand bassin se livrait à cette prise de contact quand il rentrait de la piscine de démonstration et avait donc été absent pendant un certain temps.
Quiconque possède des dauphins captifs peut faire de telles observations sous différentes formes et la littérature scientifique en fait mention.
On peut surprendre aussi de telles « conversations » de la part d’animaux en liberté. Les chercheurs californiens Evans et Dreher témoignent d’un cas remarquable qui s’est produit alors qu’ils étaient à l’ancre, à l’entrée du lagon de Scammon, en Californie.
Ils purent observer le comportement et en enregistrer les bruits d’un groupe de Tursiops qui, ne se doutant de rien, s’approchèrent d’une barrière et la reconnurent au moyen de leur sonar à 350 mètres de distance. Aussitôt le groupe s’arrêta.
Après une longue hésitation pendant laquelle les observateurs humains entendirent des sifflements, un des dauphins partit en reconnaissance, puis retourna parmi ses compagnons et fit manifestement son rapport.
Il s’ensuivit une nouvelle reconnaissance, puis une « discussion » avec des sifflements en sourdine, avant que les animaux ne passent la barrière.
Lanq et Smith ont tenté une intéressante expérience.
Ils ont installé deux dauphins souffleurs, une femelle, Doris et un mâle, Dash dans deux bassins séparés où les animaux ne pouvaient pas se voir et ils n’étaient reliés que par un téléphone.

Doris et Dash
Les chercheurs pouvaient, à l’aide d’un interrupteur, couper la ligne et empêcher chaque dauphin d’entendre son partenaire, tout en continuant à enregistrer sur bande magnétique leurs émissions sonores.
Au moment de rétablir la communication téléphonique, on émettait un signal spécial comme avertissement.
Au début de l’expérience, c’était surtout Dash qui cherchait à se rendre compte d’où provenait ce bruit. Pendant que le téléphone était en fonction, les dauphins » parlaient » sept fois plus que quand la ligne était coupée. Leurs phrases étaient courtes, quelques secondes au maximum.
Le biologiste John Lilly m’a relaté les conversations auxquelles il a assisté. Il m’a affirmé que les dauphins sont polis au point de ne jamais s’interrompre et de laisser leur interlocuteur terminer ce qu’il avait à dire.
(…)
Le problème qui se pose est de savoir dans quelle mesure les animaux peuvent se communiquer de réelles informations, ou si ce n’est là qu’une illusion. Le Docteur Bastian, de Californie, a essayé d’élucider cette question. Il a utilisé deux souffleurs qui ne pouvaient pas se voir, mais bien s’entendre.
Il apprit aux animaux appuyer sur un bouton lors de l’apparition d’un signal lumineux continu, et sur un deuxième bouton quand apparaissait une lampe clignotante.
Plus tard, l’un des animaux devait appuyer la premier sur les boutons, avant que l’autre puisse le faire à son tour. Le dressage réussit.
Dans une troisième phase, l’animal qui devait pousser le premier sur les boutons ne reçut plus de signal lumineux : de plus, il ne pouvait pas voir le signal destiné à son confrère.
Le résultat étonnant de cette expérience est que l’animal privé de la vue de tout signal, appuyait néanmoins sur le bouton convenable dans 85% des cas, à condition de pouvoir entendre son partenaire : dés que toute communication auditive était supprimée entre les deux dauphins, l’expérience échouait.

L’expérience de Bastian
L’ingénieur Van de Ree, de l’université technique de Delft, a répété les expériences de Bastian, avec un équipement plus perfectionné et des méthodes d’analyse plus raffinées: en outre, il a amélioré les processus de dressage.
L’animal qui ne pouvait pas voir le signal lumineux a été aveuglé complètement par un masque, pour exclure en toute certitude sa capacité de voir le signal.
D’autre part, l’autre animal, qui devait transmettre l’ordre qu’il avait reçu visuellement, n’était autorisé à presser le bouton qu’à la condition que son partenaire aveuglé ait lui même réagi correctement.
Au cours de la dernière série d’expériences selon ce schéma, sur 250
tentatives, 99% donnèrent un résultat positif !
Comme ultime perfectionnement de cette technique expérimentale, les sons émis par le dauphin qui « commandait » les actes de son partenaire étaient captés au moyen d’un hydrophone très sensible attaché à son museau par une ventouse adhésive: de la sorte, l’émission même du signal acoustique était enregistrée avant d’avoir subi éventuellement une distorsion dans l’eau : simultanément, les actes des dauphins étaient enregistrés visuellement sur un enregistreur vidéo.
A notre surprise, il apparut que les ordres « appuyez sur le bouton droit » et « appuyez sur la bouton gauche » n’étaient pas différents, statistiquement parlant.
Seule, la position du dauphin qui donnait ces ordres différait dans les deux cas. Cette position différente entraîne, semble-t-il, une différence dans la qualité du son émis, en sorte que l’animal qui écoute peut y trouver une information.
Il a été confirmé, comme le pensait Bastian, que les sifflements n’ont probablement aucune signification dans ce contexte. Ce sont les trains de clicks qui ont ici une importance primordiale pour le sens de l’information et pour la réussite des tentatives de communication ».
(…)
Extrait de « Extraordinaires dauphins » par le Dr W.H. Dudok Van Heel, Directeur de recherches du Delphinarium de Hardewijk
Rossel édition 1974
On notera par ailleurs que le delphinarium de Hardewijk est également l’éditeur de l’ouvrage « Sensory Abilities of Cetaceans: Laboratory and Field Evidence » sous la direction de Jeanette A. Thomas et Ronald Kastelein (Harderwijk Dolfinarium) dans la collection NATO ASI Series (Series A : Life sciences Vol.196), lequel ouvrage reprend les recherches sur le langage delphinien de Vladimir Markov.
E. Les recherches continuent !
Les recherches de Markov et de ses collaborateurs datent déjà du siècle dernier (1990) et se sont d’ailleurs interrompues avec la chute de l’empire soviétique, l’armée cessant de financer de tels travaux.
(Note 2015 : il est possible que ces recherches aient été reprises par Vladimir Poutine dans le même contexte militaire).
Depuis lors, la technologie a singulièrement évolué et il nous est possible, ainsi que l’expliquait déjà Jim Nollman dans un courrier daté de 2001 de déterminer l’empreinte vocale de chaque cétacé par des moyens purement informatiques.
Ce nouveau pas est essentiel car il permet désormais d’identifier le locuteur dans un groupe donné et de mettre en rapport les comportements observés et les vocalisations en milieu naturel, sans recourir à une quelconque capture.
Plusieurs études ont été menées sur cette base par le Professeur V. M. Bel’kovich (Académie des Sciences de Russie) à propos du langage des bélugas en Mer Blanche. Ces recherches confirment l’existence de structures phonétiques stables (« voyelles », clics, sifflements, etc.) et de l’usage intensif de schémas écholocatoires dans le discours de ces cétacés, qui pourraient être des images.
(Pour en savoir plus sur les bélugas)
Nous ne développerons cependant pas dans ce chapitre les passionnantes recherches entreprises à propos de la communication chez les orques, les baleines, les cachalots ou même chez d’autres espèces de dauphins que le Tursiops truncatus.
Les cultures et les mode de vie de ces peuples cétacés mériteraient à vrai dire des articles entiers. Les seules recherches consacrées au dauphin sont déjà abondantes et se sont développé un peu partout dans le monde dans les années 70 et 80.
Evoquons par exemple le fascinant défi qu’avait lancé l’équipe du Dr Ken Marten (Earth Trust-Projet Delphis) au Sea Life Park à Hawaï sous le nom de Project Delphis.
Avant dix ans, déclaraient-ils, nous dialoguerons librement avec les dauphins.
Le langage mis au pont par cette équipe était composé de sifflements électroniques, que les dauphins apprenaient très rapidement et dont chacun désignait un objet. Faute de crédits – et nous y reviendrons – ces recherches ont été abandonnées.
De leur côté, le psychologue James Ralston et l’informaticien Humphrey Williams ont découvert que les jeunes dauphins développent leur propre signature sifflée entre l’âge de deux mois et d’un an. Ces sifflements resteront inchangés pour la durée entière de la vie de l’animal. Au-delà de leur seule fonction nominative, certains des sifflements du dauphin apparaissent comme de fidèles reproductions de ceux de leurs compagnons et servent à interpeller les autres par leur nom.
La signature sifflée des dauphins en tant que signal complexe ont également fait l’objet de recherches approfondies de la part de Victor P. Janik, de l’Université de Saint André en Ecosse.
En avril 2000, ces recherches ont confirmé que le sifflement est un nom propre à part entière. Les travaux se poursuivent aujourd’hui parmi les dauphins de Shark Bay en Australie.
Aux USA, H. Harley, de la Division of Social Sciences, New Collège of US.A. (Sarasota, Floride) a mené le même type de recherches avec la collaboration contrainte des dauphins du Walt Disney World EPCOT Center.
De son côté, Peter Tyack (Woods Hole Oceanographic Institute) a travaillé aux côtés de David Staelin, professeur d’ingénierie électronique au M.I.T., afin de développer un logiciel d’ordinateur capable de détecter les « matrices sonores » et les signaux répétitifs parmi le concert de couinements, piaulements et autres miaulements émis par les dauphins. Une recherche similaire est menée par l’Université de Singapore (Dolphin Study Group).
Avec de tels outils, les chercheurs espèrent en apprendre davantage sur la fonction précise des sifflements.
L’Institut für Verhaltensbiologie de la Freie Universität de Berlin se préoccupe, quant à lui, des contours sonores des sifflements émis par les dauphins, en fonction de leur comportement.
P. Evans et divers chercheurs du département de Zoologie de l’université de Milan et de l’Université d’Oxford sont en train de comparer les comportements sonores du dauphin de Risso dans les eaux écossaises et dans la mer Méditerranée et cherchent à mettre à jour les dialectes respectifs de ces deux espèces.
Le Dolphin Reef Southern Beach de Eïlat en Israël, a travaillé sur les réponses données par les dauphins à certains types de sifflements artificiels. Ces recherches ont été menées en collaboration avec l’Institut fur Verhaltensbiologie de Berlin.
Il va de soi que nous ne pouvons encourager ici les recherches menées en bassin, non seulement pour des raisons éthiques mais aussi parce qu’on ne peut attendre de dauphins captifs un comportement normal ni une connaissance fine de la langue de leur tribu.
Enfin, parmi les recherches les plus passionnantes, figurent celles que mènent Denise Herzing et son Wild Dolphin Project. Sont étudiées actuellement (2012) les vocalisations des dauphins tachetés libres de manière intensive, à l’aide d’un outillage
informatique nommé « Cetacean Hearing and Telemetry (CHAT).
La relation affective intense qui relie cette chercheuse à ses « dauphins d’étude » depuis 30 ans constitue sans nul doute un gage supplémentaire de réussite.
II. Voir et parler, une même fonction

Champ perceptif et rayon sonore chez le Tursiops
Lorsque vous sortez dans le jardin et regardez une fleur, c’est le soleil qui, se reflétant sur les pétales, vous renvoie la couleur et la forme de la plante observée. Notre oeil est donc relativement passif et dépendant de la lumière solaire.
Dans l’univers obscur des fonds marins, en revanche, c’est au dauphin que revient la tâche de «jouer le soleil». A lui d’émettre des sons et d’en recevoir l’écho, exactement comme si nous sortions une lampe de poche de notre sac et que nous en éclairions le paysage nocturne.
Mais ces même sons lui servent aussi à communiquer !
On conçoit qu’une telle confusion des rôles ne reste pas sans conséquence sur l’originalité profonde de cette « langue » qui tout à la
fois perçoit les choses et les désigne !
Toute la question de sa nature se situe d’ailleurs à ce niveau : s’agit-il d’un parler d‘images holophoniques, directement copiées des formes du monde réel et transcrites en fronts d’ondes (CymaGlyphes de John Stuart Reid et Jack Kassewitz) ou bien d’un véritable langage tel que nous l’entendons, doté de vocabulaire et de syntaxe (théorie des chercheurs russes) ?
Il semble bien que ce soit les deux et que les deux approches se complètent. En quelque sorte, Vladimir Markov aurait mis en lumière l’organisation syntaxique des CymaGlyphes.

Parole de Globicéphale 1998
A. Echolocation et visiaudition
Les dauphins ne disposent pas de cordes vocales. Afin de conserver l’oxygène lors des apnées, leurs émissions sonores ne se font qu’en circuit fermé, sans dégagement de bulles d’air. De part et d’autre du conduit nasal, qui s’ouvre sur l’évent, on découvre trois paires de sacs aériens de formes et de tailles diverses.
Le cétacé contrôle leur débit avec la même aisance que nous pourrions contrôler l’ouverture d’un ballon gonflable en relâchant doucement la pression des doigts. Le passage de l’air d’un sac à l’autre détermine le son produit.
Les clicks (mais aussi toute la gamme extraordinaire que le dauphin émet par le moyen de ses sacs nasaux) sont ensuite réfléchis par la parabole crânienne située derrière les sacs aériens.

Concentrés par la « loupe » graisseuse du melon puis projetés en un seul faisceau comme la lumière d’une lampe de poche, les clicks heurtent l’objet et puis reviennent vers le dauphin. Ce dernier en réceptionne l’écho avec la pointe de sa mâchoire inférieure dont les os, très sensibles, transportent les vibrations sonores jusqu’aux oreilles internes.

La même parabole crânienne joue à ce moment le rôle d’antenne et non plus d’amplificateur et le savant travail de décodage peut alors commencer dans le cerveau du dauphin.
Imaginons à présent un exemple concret, afin de nous rendre compte de ce que peut être la « vision acoustique » fournie par ce mécanisme de l’écholocation.
Un dauphin nage la nuit, sous dix mètres de fonds. De temps à autre, il émet une décharge de clics à basse fréquence, qui portent à une centaine de mètres et ne rendent compte que de la présence des masses importantes.
Ces « burst pulsed sounds » extrêmement brefs (moins d’une seconde d’émission continue) ne sont pas des sons purs mais des « bruits », d’inextricables petits paquets d’ondes situés sur des fréquences de 120 à 130 Khz et d’une puissance frisant parfois les 220 décibels.
Ils retentissent sous l’eau comme une grêle de minuscules coups secs et nets enchaînés l’un à l’autre en de courtes séquences. Concentrés par le melon, les clicks sont émis sous forme d’un large faisceau, qui balaie par intermittence le sol sablonneux à la façon d’un projecteur. Les informations reçues, assez grossières, concernent l’aspect du fond marin ou une masse importante, bateau ou autre cétacé.
Supposons à présent qu’un poisson soit repéré dans ce champ de vision « stroboscopique ».
Puisqu’il fait nuit, l’oeil ne peut confirmer l’image en mode visuel. Lorsque la chasse commence, le dauphin resserre alors le rayon de son biosonar et le dédouble en deux faisceaux.
Plus précis, mieux ciblés les trains de clicks bombardent le poisson sous tous ses angles et peuvent même pénétrer dans son corps en renvoyant l’image de ses organes internes. Les deux trains de clicks sont produits presque simultanément, l’un à 20° à gauche de la ligne du rostre et l’autre à 20° sur la droite. Les deux rayons se chevauchent au point focal (0°) et fournissent une « visiaudition » de type binoculaire.
Un intervalle de 80 millièmes de seconde sépare l’émission de chacun des faisceaux, de sorte qu’en calculant le léger retard d’un écho par rapport à l’autre, le dauphin peut estimer la profondeur de champ et la distance qui le sépare de chaque élément de l’objet observé.
Se rapprochant de sa proie à toute vitesse, le dauphin n’a de cesse que de conserver le contact avec elle et multiplie la fréquence et l’intensité de ses trains de clicks, comme pour maintenir le « projecteur » allumé presque en continu.
Les ondes à haute fréquence ont une portée plus courte mais fournissent en revanche une bien meilleure définition des détails.
En nageant, le dauphin opère un mouvement de balayage avec la tête avant d’obtenir une image complète de sa cible, que ses organes visuels conforteront par ailleurs. S’il veut obtenir davantage de détails encore sur son contenu, le dauphin la bombardera alors sa cible à bout portant, d’un faisceau de clicks aussi fin et précis qu’un rayon laser.
Celui-ci pénètre la matière et en estime la densité avec une incroyable précision : la nature d’un métal (zinc plutôt que cuivre) ou des variations de l’épaisseur d’un tube de l’ordre d’un millième de millimètres sont alors parfaitement perçus par cette échographie biologique.
B. Echogramme et trasmission d’images

L’écholocation est aussi un langage
Selon H.Jerison (1), l’image fournie par les deux échos-retour – avec un décalage de quelques millièmes de seconde – évoquerait celle qui naît du mouvement d’un pixel sur un écran de télévision. Le parcours rapide d’un minuscule éclair lumineux d’intensité et de fréquence variable imprime dans la rétine du spectateur une série de schémas successifs à très brefs intervalles.
L’effet de rémanence reconstruit alors une image spatiale en mouvement parfaitement lisible au seul départ de ce bombardement photonique pointilliste.
De la même manière, le dauphin ne perçoit du poisson qu’un réseau complexes d’échos ponctuels successifs que son cerveau apprend à lire.
La même rémanence rétinienne qui nous permet de regarder un film permet au cétacé de garder en mémoire cette suite de crépitements qui lui revient. Chaque « échogramme » est reconnaissable et le dauphin le traduit aussitôt en éléments d’une forme globale.
Enfin, les zones cérébrales supérieures dévolues à la vision et à l’audition traitent ces données et l’image nette d’un mulet rayé surgit pour le dauphin dans les ténèbres aquatiques.
La mémoire du regard complète sans doute l’information, mais on peut parfaitement se figurer l’image que le dauphin perçoit : en noir et blanc et en relief, exposant ses organes et les battements de ses vaisseaux sanguins, le poisson pâle traîne derrière lui une aura d’étincelles résultant de ses propres productions acoustiques et des multiples turbulences dues à ses déplacements.

Imaginons maintenant qu’un second dauphin se trouve dans les parages. Il circule assez loin, le dos tourné au premier et regardant autre chose. Dès que le dauphin A aura ciblé son poisson et qu’il en recueillera l’écho, le dauphin B, même à dix mètres, percevra lui aussi l’image parfaite du même mulet rayé, comme s’il venait de focaliser lui- même cette proie !
Rien de plus normal, puisque l’écho se diffuse largement en milieu aquatique. Une partie en sera reçue par le menton du dauphin A tandis que le reste fera vibrer tout le corps du dauphin B.
A ce stade, on le comprend, l’écholocation s’apparente davantage à la transmission de pensée qu’à une simple technique de repérage et de chasse.
Penner et Kadane (1980) ont ainsi pu apporter les preuves expérimentales de ce que l’écho de chaque objet, chaque personne ou chaque animal est parfaitement mémorisé et même reproductible « à vide ».
Hammer et Au (1980), de leur côté, ont clairement observé que les dauphins se livraient à l’écholocation en l’absence de toute cible, procédant de cette manière à l’énonciation d’un mot et non plus à sa perception.
Nous verrons plus loin comment les audiogrammes perçus lors du retour d’écho représentent une sorte de « nom naturel » et constituent peut-être le fondement du vocabulaire dauphin. Sans doute n’ont-ils plus alors leur précision initiale. L’usage raccourcit tous les mots et certainement les images aussi. Au lieu d’une icône spectrographique complète, le dauphin ne lance plus que l’amorce du son ou ses contours. L’observation nous apprend d’ailleurs qu’un tel usage est courant chez les dauphins lorsqu’ils sifflent leur « signature » (voir ci-dessous). Souvent, celle- ci n’est plus émise qu’en version courte, abrégée, esquissée rapidement.
C. Sons purs et signature sifflée

Chaque dauphin porte un nom
Outre ces brefs sons pulsés, émis sur une large fréquence et de durée très courte, les dauphins produisent également d’autres sons à l’aide de leurs mêmes sacs aériens.
L’un des sons les plus connus est le fameux « sifflement » que le dauphin peut émettre avec puissance sous l’eau et parfois hors de l’eau, toujours par l’évent.
Il s’agit cette fois d’un signal non-pulsé, continu et modulé dans des fréquences suraiguës, le plus souvent imperceptibles à l’audition humaine.
A l’analyse spectrographique, on constate que ces signaux sont des sons purs, non mélangés, qui peuvent durer de 0,5 à deux secondes.
Puisqu’il est plus aigu, un son sifflé porte naturellement plus loin que les clics ordinaires, ce qui permet ainsi aux membres éloignés d’un groupe de rester en contact.
Le répertoire sifflé d’un dauphin Tursiops est émaillé d’un son très particulier qui n’appartient qu’à lui, la signature sifflée (signature whistle).

Lorsqu’ils quittent leur famille à la fin de l’adolescence, les jeunes mâles peuvent rencontrer au cours de leurs voyages un groupe de femelles issues de la même tribu (« pod ») qu’eux. Pour éviter toute relation incestueuse, chacun décline alors son identité. Ou bien encore, lorsque deux frères se retrouvent au terme de longues errances et qu’ils se reconnaissent comme tels, jamais aucun combat, duel ou rivalité amoureuse n’adviendra entre eux deux.
Selon l’intensité et la modulation de ce son – émis vingt ou trente fois à la seconde – il exprime également son état émotionnel ou signale simplement sa propre présence à ses compagnons. « Je suis là… je suis ici… Moi ici… » se lancent sans cesse les dauphins qui nagent, chacun égrenant son nom aux quatre coins de la troupe.
Plus la nuit tombe, moins on y voit, et plus les dauphins s’interpellent alors et dialoguent sous les eaux. Mais ce système, continue le Dr R.Wells, n’est pas le seul en action chez les dauphins. Dans le cas d’une conversation d’individu à individu, un dauphin fera suivre son propre sifflement par celui de son interlocuteur, mais selon des règles subtiles. Par exemple, l’ordre de l’énoncé tiendra compte de la position hiérarchique de celui qui s’exprime.
Par ailleurs, on observe que les dauphins effrayés ou échoués émettent sur une fréquence souvent accélérée, qui n’est pas sans rappeler le débit altéré d’un humain en détresse. Dès qu’un dauphin se met à émettre un signal en boucle, les autres s’agitent ou le rejoignent.
Enfin, lors des naissances, toutes les femelles rassemblées qui entourent la parturiente, sifflent leurs propres noms en boucle, sans cesse, comme si elles chantaient…
On ajoutera pour conclure que d’autres études, plus récentes, ont révélé que le répertoire des sifflements ne se limitait pas aux signatures sifflées. D’autres modulations existent, qui n’ont pas encore fait l’objet d’un recensement systématique ni n’ont encore été mis en corrélation directe avec les comportements observés. Elles ne s’entendent en effet qu’en milieu naturel.
Pour Denise L.Herzing (Florida Atlantic Unversity) qui travaille depuis des années avec les dauphins tachetés libres vivant aux Bahamas , la notion de signature sifflée fixe telle que décrite par Randy Wells mais aussi par Sayligh et al (1990) ne doit pas occulter le fait que cette « signature » sert en fait à d’autres usages que la simple signalisation du nom. Denise Herzing observe en effet qu’il existe un véritable répertoire de sifflements nuancés selon les circonstances , sans parler de l’ajout des bulles d’air comme « accents » du discours, et que de façon générale, ces sifflements sont produits :
– durant la période d’excitation précédant la chasse nocturne chez le «spinner dolphin»
– durant les jeux sur la lame d’étrave des navires (bow riding) chez le dauphin commun
– durant la période d’alimentation des globicéphales
– durant les activités menées en coopération par plusieurs individus chez le Tursiops
– lors des réunions mère-enfant chez le Tursiops.
Note 2015 : les recherches sur la signature sifflées ont beaucoup progressé depuis ces découvertes. Il apparaît par exemple que les membres d’un Trio de mâles modifieront subtilement leur propre signature sifflée pour qu’elle ressemble à celle de leurs compagnons : le trio finit donc par n’avoir qu’un seul nom !
D. Infrasons et autres productions sonores
Un dauphin adulte bien exercé est capable d’émettre en une seule fois quatre sons différents, issus de quatre sources distinctes.
C’est apparemment là tout l’art de la langue dauphin : parvenir à combiner, comme sur des grilles harmoniques, deux, trois voire quatre couches superposées de clicks, sifflements et autres productions sonres encore plus insolites. Seuls les plus âgés y parviennent.
En langage courant, deux sources sonores – l’une pour le sifflement, l’autre pour le click – suffisent à composer des phrases complexes. En outre, un jeu d’échos et de silences permet encore de moduler les phrases, comme on le verra plus loin.
Richard C.Connor a tenté de classer ces sons en (au moins) 8 catégories :
1. Barks (aboiements)
2. Donkey’s bray (braiments d’âne)
3. Squeaks (cris pulsés d’intensité croissante)
4. Squawks (cris rauques)
5. Grunts (grincements, grognements)
6. Mews (miaulements)
7. Moans (gémissements)
8. Yelps (jappements)
Denise L.Herzing y ajoute encore les vocalisations suivantes :
– Excitment vocalization :
Vocalisation d’excitation. Sons pulsés avec signature sifflée en surimpression, 2-30secondes de durée, 4à 18 kHz en puissance sonore, généralement accompagnées
d’émission de bulles et produit dans des circonstances de détresse ou de grand énervement.
– Genital buzz :
Clicks de haute fréquence répété envoyé rapidement sur la zone génitale d’une compagne ou d’un compagnon. 1,2 Khz- 2,5 kHz 8-2000 clicks par secondes. De 6 à 20 secondes de durée. Utilisé lors des interactions amoureuses.
– Razor buzz :
Train de clicks écholocatoires de 2, 0 à 6 kHz. 200 clicks par seconde, maintenus pendant plusieurs minutes. Le son est typiquement celui d’un rasoir électrique ! Utilisé lorsque les dauphins se regroupent en faisceaux, couchés sur le fonds sableux.
– Echolocation avec rostre dans le sable :
2, 0 à 6 kHz, 400 à 500 clicks par seconde, durée brève de deux à trois secondes. Utilisé lors du foraging
– Echolocation avec trilles «recouvrantes» et sifflements «surélevés»
2, 0 à 6 kHz , 8 à 100 clicks par seconde, trilles répétées sous 5 kHz, sifflements de 4,8 à 16 kHz. Utilisé par les dauphins Tursiops des Bahamas lors de l’exploration des rochers et des bancs de récifs. Durée brève de deux à trois secondes.
Les dauphins peuvent encore émettre quantité d’autres sons qui s’apparentent à des cris ou à des ordres brefs, et qui sont toujours produits dans un contexte émotionnel intense. Lorsqu’un dauphin se met en colère, il agite généralement la tête en produisant un violent « crack » explosif.
Lorsqu’il appelle une jeune delphine de façon impérative, il émet plutôt une sorte de « pops », qui semble vouloir dire « Viens ici ! ».
E. Langage gestuel chez les dauphins Tursiops

Différents « styles de nage » .
Selon qu’il montre son ventre ou son dos, qu’il se tourne ventre vers le ciel ou pointe son rostre dans le sable, qu’il place son corps en S ou nage d’une certaine façon, le dauphin exprime mille nuances dans le même temps qu’il vocalise.
La richesse expressive de cette gestuelle est immense, compte tenu des possibilités de mouvement dans un milieu en apesanteur. Par ailleurs, l’émission de bulles savamment contrôlées permet également aux dauphins de nuancer leur discours de façon subtile.
En d’autres termes, de la même manière que nos gestes ou nos mimiques faciales ajoutent du sens à nos discours verbaux, et en constituent même le complément indispensable, les dauphins modulent leurs expressions verbales par une gestuelle très fine et précise, dont nous ne connaissons malheureusement pas les codes.
D’autres études s’appliquent d’ailleurs à combiner les données acoustiques avec les données visuelles : quel est le son qu’émet un dauphin lorsqu’il se place dans telle position ou exécute tel mouvement ? Y a-t-il une corrélation entre ces éléments ?
Ces observations sont les premiers pas du long et difficile chemin qui nous mènera un jour au décryptage complet – avec l’aide des dauphins eux-mêmes, on le suppose – d’une pensée fondamentalement « autre » s’épanouissant dans un milieu autre.
D’ores et déjà, la prodigieuse complexité de la communication des cétacés qui combinent sons, mouvements, émissions de bulles et positions relatives des corps a de quoi nous donner le vertige.
Il faut lire à ce propos une fascinante étude intitulée « Nouvel inventaire du comportement du grand dauphin (Tursiops truncatus) Approche comparative des comportements des dauphins grégaires, solitaires et familiers » par M.Müller, H.Bouytière , ACF Weaver et N.Candelon.
Publiée en 1998 par le GREMMS et la revue « Vie et Milieu », Vol.48 N°2 – Juin 1998, cette recherche regroupe un vaste ensemble d’attitudes physiques et de mouvements propres aux dauphins, par le moyen desquels ils s’expriment et dont voici quelques uns parmi les plus familiers ou les plus intrigants…
Comportements aériens
* Bond hors de l’eau (Bow)
Saut hors de l’eau sur une distance ne dépassant pas la longueur du corps. Le pédoncule caudale est en l’air pendant que la partie antérieure plonge. Soit le pédoncule caudale, soit la partie antérieure, est fixe tandis que l’autre est mobile.
* Tête hors de l’eau (Head up)
La partie avant du corps est hors de l’eau dans une position oblique. Le dauphin peut rester temporairement immobile mais le corps est franchement arqué.
* Bond sur le côté (lateral bow)
Le dauphin bondit hors de l’eau sur le côté ou bondit le corps droit et pivote lorsqu’il est hors de l’eau.
* Eclaboussement de côté
Le dauphin s’élève en partie hors de l’eau et se laisse retomber bruyamment à plat.
* Eclaboussement arrière (Back Breaching)
Le dauphin élève la partie avant hors de l’eau et se laisse retomber bruyamment en arrière, en éclaboussant.
* Saut en long (Leap)
Le dauphin saute hors de l’eau en position normale sur une distance nettement plus grande que la longueur de son corps.
* Saut sur le dos (inverted leap)
le dauphin saute hors de l’eau en position inverse, aileron dorsal vers le bas.
* Saut sur le côté (lateral leap)
Le dauphin saute et retombe sur le côté
* Saut à travers la vague (Wave-leaping)
Réalisation d’un saut depuis l’arrière d’une vague par un dauphin qui se déplace dans le sens opposé à celui de la vague.

Le langage des sauts
Flexions (aches)
Corps tendu, étiré, courbé, chandelle verticale, etc.
Comportements de contact
Parmi ces comportements, citons la traîne d’un objet (Kelp dragging) : un dauphin traîne en surface un objet – le plus souvent une algue – accroché à l’avant de la nageoire dorsale ou à une autre partie du corps.) mais aussi le coup de rostre, le coup de rostre vers le bas, le frottement, la prise en bouche, etc.
Mouvements de tête
* Menton en l’air (chinup)
Brèves expositions du rostre hors de l’eau selon des angles variables par une élévation de la tête .
* Lancer de poisons (Fish toss)
Un dauphin tenant un poisson entre ses mâchoires, émerge la tête hors de l’eau. Le poisson est envoyé sur le côté ou en arrière par un redressement brusque de la tête.
* Coup de rostre vers le haut (Beak up)
Le dauphin stationnaire, en positon normale, effectue des mouvements de rostres vers le haut, souvent pour toucher un objet.
* Oeillade
Brève sortie d’un oeil hors de l’eau par un dauphin stationnaire ou se déplaçant en position normale par une rotation sur le côté ou en abaissant le rostre vers le bas.
* Salut de la tête (Head nod)
Un mouvement de tête de haut en bas hors de l’eau ou sous l’eau par un dauphin en position normale.
* Salut aérien (Airbone headnod)
Le dauphin effectue un signe de tête pendant un saut , tout son corps étant hors de l’eau , juste avant la rentrée dans l’eau. (…)
* Toupie (Head turn)
Dans une position verticale, le dauphin émerge la tête ou le rostre hors de l’eau et pivote sur lui-même une ou plusieurs fois. Comportement également observé sous l’eau.
* Bouche ouverte (Mouth open)
Le dauphin ouvre la bouche légèrement et lentement, souvent avec un mouvement coordonné de la tête. Ce comportement est souvent effectué sous l’eau mais parfois la tête hors de l’eau.
Mouvements des nageoires pectorales (Pectoral fin movements)
* Une pectorale hors de l’eau (Pectoral extension)
Exposition à la surface d’une nageoire pectorale par un dauphin qui se tient sur le côté. L’angle entre la pectorale et l’axe longitudinal est aigu.
* Les pectorales parallèles hors de l’eau (Pectoral parallel extension)
Exposition à la surface des deux nageoires pectorales par un dauphin qui est sur le dos, stationnaire ou en mouvement.
* Flexion d’une pectorale (Pectoral flex)
Mouvements rapides, du haut vers le bas, d’une nageoire pectorale exposée à la surface.
Respiration
* Ebrouement (Chuffing)
Emission d’air brusque et très bruyante. Un petit cône de vapeur condensée s’élève à un mètre environ au-dessus de la tête de l’animal.
* Expulsion chuintante (Squeeze breath)
Expulsion d’air par l’évent , brève et sonore, rappelant le bruit de l’air s’échappant d’un ballon d’enfant qui se dégonfle, avec très peu de vapeur condensée visible.
* Emission d’une grosse bulle (One large bubble)
Emission sous-marine d’air par l’évent formant une ou plusieurs larges bulles à la surface. (…)
Claquements (Slaps)
* Claquements du menton (Chin slap)
Elévation et abaissement du rostre qui frappe bruyamment la surface de l’eau. A faible intensité, c’est uniquement le rostre qui est utilisé. A forte intensité, c’est toute la partie avant du corps qui est élevée hors de l’eau et rabattue violemment.
* Claquements de la nageoire dorsale (dorsal fin slap)
Le dauphin effectue des balancements des deux côtés de l’axe longitudinal depuis une position normale. La nageoire dorsale vient heurter la surface de l’eau à plat.
* Claquements de la tête sur le côté (Head slap)
Le côté de la tête du dauphin frappe bruyamment la surface de l’eau. L’animal peut marquer un temps d’arrêt et étendre ses nageoires pectorales avant de se laisser retomber.
* Claquement de la tête en avant (Forward head slap)
Ce comportement est observé lors d’un déplacement en avant, le dauphin sortant seulement la tête hors de l’eau en la claquant fortement contre la surface.
* Claquement de la tête en arrière (Back head slap)
Puissant contact de la région de l’évent à plat avec la surface par un jeté violent de la tête en arrière. Le dauphin, d’abord en position normale, sort la tête hors de l’eau pour atteindre progressivement une position verticale.
* Claquement de la pectorale (Pectoral fin slap)
Contact rapide, bruyant et répétitif entre la nageoire pectorale et la surface de l’eau, effectué par un dauphin couché sur le côté.
* Claquement de la queue (Tail slap)
Contact bruyant de la partie arrière du corps à plat avec la surface de l’eau. A faible intensité, seule la caudale s’élève hors de l’eau. A forte intensité, le pédoncule caudal entier est élevé et frappe la surface. La partie avant est souvent sous l’eau et les claquements peuvent être répétitifs.
* Claquement de la queue à l’envers (Inverted tail slap)
La partie dorsale de la caudale d’un dauphin sur le dos frappe à plat la surface de l’eau . Ce mouvement peut être répété. Il est observé chez un dauphin immobile ou en mouvement.
* Claquements en série (Motorboating)
Série de claquements rapides et répétitifs de la queue pendant le déplacement d’un dauphin en position normale ou sur le dos.
Comportement stationnaire (Stationary behavior)
* Flottaison (Floating)
Situation d’un dauphin immobile à la surface en position normale. La région de l’évent est très bombée car la tête est abaissée. Seul le sommet de la nageoire dorsal est visible.
* Flottaison à l’envers (Ventral floating)
Situation d’un dauphin immobile à la surface en position normale. L’animal apparaît voûté, avec seulement la tête et les pointes de la caudale hors de l’eau.
* Roulis atténué (Lean)
Rotations légères sur le côté sans laisser émerger l’oeil
* Radeau (Rafting)
Situation d’un dauphin étendu immobile en surface.
* Mouvement de détente (Resting Mouvement)
A la sortie d’une position de radeau, le dauphin fait émerger lentement la partie avant du corps jusqu’à avoir le menton en l’air tandis que le pédoncule caudale est redressé. L’animal expire de l’air dès son arrivée en surface ou bien juste avant l’immersion. (…)
* Roulis accentué (Roll)
Rotations accentuées autour de l’axe longitudinal par un dauphin en position normale. La rotation a souvent lieu d’un seul côté. La nageoire dorsale s’approche plusieurs fois de la surface de l’eau sans toutefois la toucher. (…)
* Position verticale (Vertical position)
Maintien dans une position verticale sous l’eau , le rostre pointant vers la surface . Cette position est souvent tenue pendant une ou plusieurs minutes.
* Emersions et submersions
Plongée arquée, immersion vers l’avant, sortie de l’eau à l’horizontale, enfoncement sur place. L’animal coule à pic d’un seul tenant, en position horizontal puis reste sous l’eau sans bouger.
Etc.
Style de nage
Nage dans la vague (Swell-riding) :
Un dauphin en position normale se laisse porter par la houle. Immobile, l’animal attend qu’une vague passe puis se laisse glisser avec elle, exposant la nageoire dorsale et le pédoncule caudal.
Nage à l’étrave d’un bateau, nage en tire-bouchon, glissade, nage en zig-zag, nage sur le dos, nage sur le côté, marsouinage , émersion bouillonnante, nage sous la surface, nage au ras de la surface, surfing sur le dos des vagues, nage ondulée, nage ventre à ventre, etc.
Cette liste n’est, rappelons-le, qu’une infime partie des milliers d’attitudes gestuelles connues et répertoriées chez le seul dauphin Tursiops.
On lira également à ce propos l’étude en anglais sur la gestuelle des dauphins d’Irlande
F. Syntaxe et vocabulaire
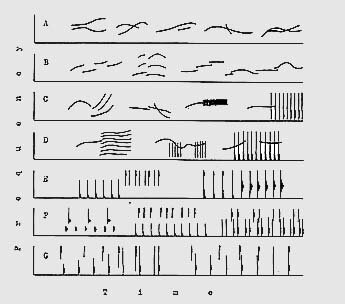
Les deux registres du sifflement (son pur) et des « trains de clics » (sons composés) se trouvent toujours associés dans les conversations entre dauphins, contrairement à l’opinion des premiers chercheurs qui pensaient que les clics n’avaient d’usage qu’en terme d’écholocation. Il s’agit en fait d’un double mode d’expression parfaitement homogène, inusité chez nous, qui fonctionne en « surimpression ». Seuls les chanteurs Mongols peuvent produire deux émissions sonores simultanées, et cette pratique n’intervient qu’à titre de recherche esthétique, jamais dans l’usage courant.
Lorsque les dauphins sifflent, leurs clics restent parfaitement distincts et audibles, puisque leur fréquence est plus basse que celle des sifflements.
Ceux-ci sont toujours composés d’éléments simples, aisément reconnaissables, qui s’articulent en structures plus complexes – on pourrait dire : multidimensionnelles – pour former des chaînes de « blocs sonores », exactement de la même manière qu’un mot s’articule à d’autres pour former des phrases. Ces grandes structures elles-mêmes se répètent en « loops » selon certains rythmes, qui semblent d’une importance extrême dans la logique même du discours.
« Lorsque l’un des quatre générateurs de sons fonctionne en régime tonal », explique à ce propos Vladimir Markov, « il produit des signaux en modulation de fréquence (des sifflements) sur une bande étroite.
En variant la direction et le degré de variation de la fréquence, un dauphin peut produire diverses structures acoustiques parfois bizarres. Lorsqu’on les analyse de plus près, on note que ces structures sont produites par des sections alternant de manière arbitraire avec une rapide augmentation ou diminution de la fréquence assortis d’autres segments dotés d’une fréquence variant peu ou lentement.
Le résultat est que la structure du signal devient une chaîne de différents éléments acoustiques et développe ainsi les possibilités de contrastes nécessaires à l’encodage d’une information. Les capacités de stockage de l’information fournie par ce système peut encore être augmentée en modifiant l’angulaire (steepness : raideur, escarpement) des contours de ces segments, en changeant les limites du domaine de fréquence, le registre (position du contour sur l’axe de fréquence) et la durée, aussi bien qu’en augmentant le nombre total d’éléments composant le signal »

Divers modules constitutifs d’un sifflement complet
* Ces phrases sifflées interagissent elles-mêmes avec divers types répertoriés de « trains de clicks » qui peuvent émis seuls comme ci-dessous :
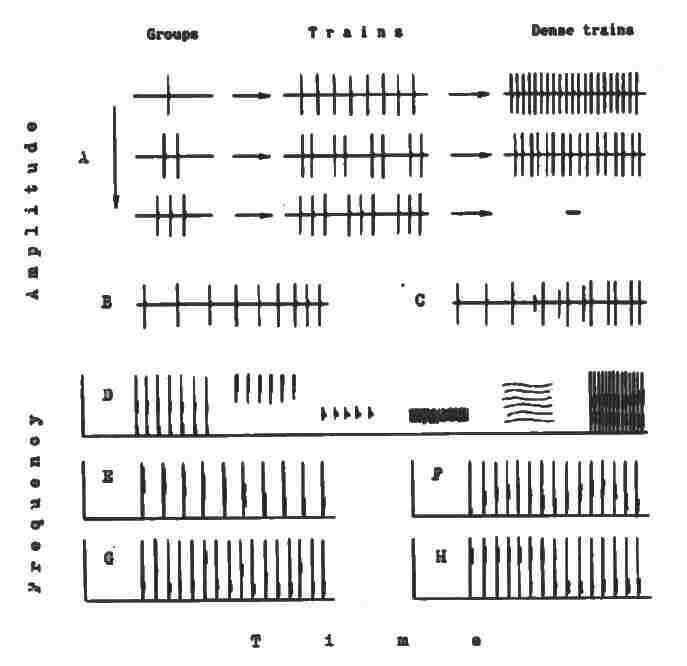
… ou en surimpression « derrière » ou « par-dessus » le son des sifflements. On notera sur le même tableau que les chaînes de sifflements peuvent se combiner uniquement entre elles ou les clicks uniquement entre eux, selon le même processus de surimpression.
Sifflements et clicks combinés en phrases
Les clicks constituant ces « trains » ne sont jamais arbitraires ni nécessairement simples : ils s’organisent eux aussi selon certains schémas convenus et peuvent à leur tour être modulés, se raccourcir, s’allonger ou se transformer en braiments, piaulements, miaulements ou sons explosifs.
Enfin, l’ensemble de ces blocs sémantiques (clicks de diverses natures, segments sifflés en combinaison) s’enrichissent de modalités subtiles : échos, vagues spectrales, crescendo et decrescendo, ralentissement ou accélération du débit, harmonisations multiples, etc.
Plus extraordinaire encore, ces éléments s’articulent en définitive en longues structures d’un ordre supérieur, comparables à des « textes » selon le mot de Markov, textes dont on a tout lieu de croire qu’ils sont porteurs de sens !
Il est clair qu’à ce stade, décrit dans le dernier chapitre de l’étude russe à grand renfort de formules mathématiques, il n’est plus question de « langue delphinienne » considérée sous un angle global mais bien d’un « dialecte des dauphins de la Mer Noire » propre aux populations locales et le seul à ce jour à avoir été étudié de façon systématique.
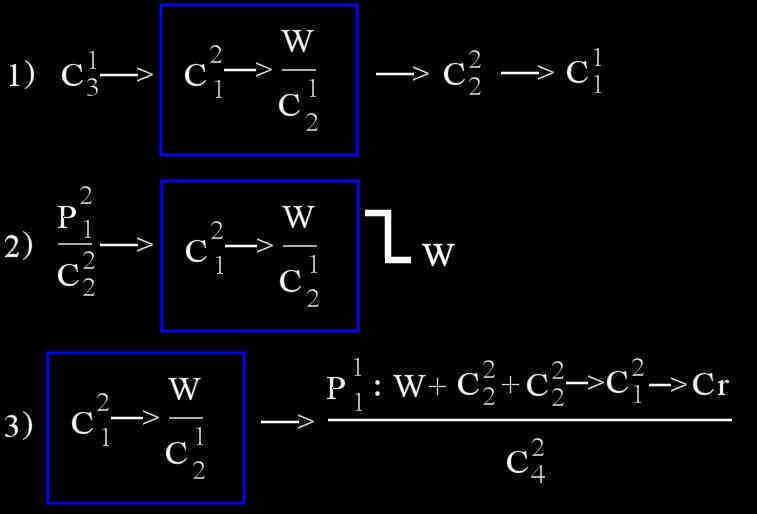
Construction syntaxique du discours delphinien
Aperçu du commentaire de Markov:
Ici, (W) vaut pour les composants tonales (sifflements) des signaux,
indépendamment de la complexité de leur structure.
(C) vaut pour les sons pulsés de la classe des clicks.
(B) appartient à la classe des « clear blows » (sons explosés).
(P) vaut pour la classe des « sons explosés prolongés ».
Les chiffres indiquent notamment l’intensité du train de clicks, etc.
Rappelons que pour obtenir ces résultats, 20 dauphins adultes ont été capturés pour l’occasion parmi lesquelles 7 mâles, 13 femelles et trois nouveaux-nés. Enregistrés soit dans des enclos en mer, soit dans des bassins, ces individus communiquaient entre eux directement ou via un canal électro-acoustique, lorsqu’ils étaient isolés. Le total des sons enregistrés sur bande magnétiques s’élevaient à plus de 300 000.
G. Icônes holophoniques

Ondelette de dauphin de l’Atlantique
Nous avons vu plus haut comment le dauphin prenait connaissance de son environnement par le moyen de l’écholocation.
A chaque objet, à chaque personne, à chaque animal, le dauphin associe rapidement l’écho correspondant et très vite, l’atténue ou l’abrège lorsqu’il le communique à son tour.
On peut donc supposer – et c’est la thèse de l’holophonie défendue par Jim Nollman – que le processus de conceptualisation et de découpage du Réel en unités discrètes sous la forme de sémantèmes a pu s’opérer chez le dauphin par le biais de ses échogrammes, suscitant de la même manière une langue ouverte et complète.
Nous imaginerons donc à titre d’hypothèse, cette fois – comment les « clicks écholocatoires » – c’est-à-dire le reflet même de la réalité externe sous le regard du sonar – peuvent donner naissance à des noms concrets, à des symboles d’objets esquissés, selon la même logique qui prévaut pour les idéogrammes chinois. Pour nous, dont le cerveau n’est pas même de décoder de telles images sonores, ces « noms » apparaissent sur l’écran d’ordinateur sous forme de cymaglyphes ou d’ondelettes.
Le rôle des sifflements est plus complexe, et nous suggérerons ici qu’ils peuvent exprimer les modalités les plus abstraites du langage dauphin. Les noms propres étant sifflés, on peut supposer qu’il s’agit de l’équivalent d’adjectifs ou de qualificatifs et que cette règle est générale.
Quant aux autres types de sons, ou à la valeur des sons polyphoniques au simple titre de maîtrise phonatoire, nous n’en savons pas assez aujourd’hui que pour leur attribuer une fonction certaine.
Imaginons, de façon grossière et certainement approximative, le processus de complexification qui pourrait s’opérer à partir d’un échogramme simple, tel que celui d’un poisson bien réel.
1. Echolocation simple
Dans un premier temps, le dauphin scanne sa cible.

Emission de clicks
2. Image-retour totale
L’écho lui revient sous forme de paquets d’ondes légèrement décalées et lui offre une photographie parfaitement lisible du poisson qu’il regarde, en l’occurrence, une femelle avec le ventre bourré d’oeufs. On peut dire qu’à ce stade, l’écho-retour est interprété en tant que « ce poisson-là » en particulier. Si le dauphin voulait informer un ami de l’existence de ce poisson, il émettrait alors l’échogramme complet, en relief et avec tous ses détails.

« Ce poisson-là »
3. Conceptualisation
Supposons maintenant que le dauphin ne veuille exprimer que le concept de poison, en tant que
« nourriture » ou « acte de se nourrir » ou comme un adjectif équivalent de « comestible ». Il enverra alors à son compagnon non pas l’écho scrupuleusement complet du poisson A mais bien une sorte d’esquisse plus rapide, convenue, presque déjà symbolisée.

4. Amplification symbolique par répétition
Le dauphin aimerait ajouter une nuance. Ce n’est pas un poisson qu’il a vu tout à l’heure, mais toute une multitude, et son excitation est grande de le faire savoir. Les échogrammes sont produits très vite en très grands nombres. Cette pluie d’images identiques devient donc, tout naturellement, le synonyme de « multitude », « banc de poissons » mais aussi du verbe « se multiplier » ou de l’adverbe « beaucoup ».

« Plein de poissons ! »
5. Complexes logiques
Le dauphin parle à une femelle. Il l’aime bien, il lui suggère une parade amoureuse et lui assure que de tous les mâles, il est le plus fertile et le mieux susceptible de lui donner un bel enfant. L’écho rapidement esquissé d’un embryon de delphineau couvert en filigrane par celui de « multitude » donne le complexe logique attendu : fertilité, descendance, vieillesse heureuse, etc. »

« Une belle descendance ! «
6. Dérivation métaphorique
La femelle lui répond qu’elle lui préfère un autre mâle, plus mûr, plus sage. Afin de se faire bien comprendre, elle associera le nom de ce vieux dauphin avec une suite de « symboles mutuellement interprétatifs ». L’image d’un aileron isolé de son contexte et marqué de cicatrices signifie beaucoup de choses dans la culture dauphin. Un adulte ainsi marqué a beaucoup voyagé, il a survécu à de nombreux dangers, il est fier, vigoureux, relativement âgé, etc.
Prise dans son large ensemble, cette icône acoustique vaut donc pour les notions voisines de « courage », « bravoure », « longs voyages » ou pour les déterminatifs associés (ancien, vieux, courageux).
Si l’accent est mis sur la blessure elle-même, que l’écho renvoyée détaillera davantage, alors la nuance obtenue sera celle de « douleur », « combat », « agression », « danger », etc.

« Un voyageur expérimenté »
7. Opérateurs modaux
Combinés avec les grilles de clicks, des sons purs sifflés n’ont cessé d’être émis durant toute cette conversation. Certains auteurs supposent que leur fonction est celle d’opérateurs modaux.
Selon le mot de Julia Kristeva – et les théories de Ken Levasseur – ils seraient des « symboles de symboles », esquissant les contours d’échogrammes oubliés, à la manière des « mots vides » de la langue chinoise.
Les sifflements n’exprimeraient plus dès lors que des concepts purs, des notions abstraites privées de tout équivalent concret, des adjectifs qualificatifs (d’où les noms propres), des verbes ou bien encore des adverbes de lieu (ici, plus loin, là-bas) de temps (hier, aujourd’hui, demain) ou de causalité (pourquoi, parce que, à cause, etc.)
Associé à l’image rapide d’un encéphale en transparence – équivalent au mot « esprit » – un sifflement particulier peut exprimer l’association, la dualité, le chiffre deux, la gémellité. Le groupe vocal signifie donc « amour » ou « amitié »

« Deux esprits ensemble »
Toutes ces opérations mentales sont parfaitement accessibles au dauphin. Les expériences d’Herman l’ont montré capable de prouesses sémantiques inouïes et nous savons qu’il domine parfaitement l’identification vocale (vocal labeling) des objets et des gens.
Le rôle du sifflement dans le contexte de l’holophonie demeure cependant mal compris. Ne perdons pas non plus de vue l’existence de tous ces sons annexes, ces braiments, miaulements et autres grondements syncopés. Souvenons-nous que le dauphin imite les sons à la manière du perroquet et qu’il émaille peut-être en liberté ses discours de bruits de moteurs, cris de mouettes ou autres aboiements de chien, comme il peut le faire en captivité.
Quoiqu’il en soit de nos connaissances actuelles, il nous est pourtant déjà possible de travailler sur des faits établis. L’écoute attentive d’enregistrements de sons delphiniens est à cet égard riche d’enseignements.
Notons aussi les récentes recherches (2003) de Mark Fischer sur les graphes des ondelettes, ainsi que celles sur les cymagyphes qui conforte de manière frappante la théorie d’un vocabulaire holophonique.
H. La logique de la langue
La question de la logique interne de la langue delphinienne et de ses catégories ne nous est actuellement pas accessible. Les expériences de Bastien et d’Evans doivent être aujourd’hui relues sous cet angle et bien entendu, toutes celles de Markov et Ostrovkaya.
La capacité de Phoenix et Akekamaï à user de grammaires inversées ou leurs tendances à privilégier le contexte circonstanciel (en quel lieu, à quel moment) au détriment du sujet (qui) et même du verbe d’action sont déjà des indicateurs troublants d’une logique interne à la langue sensiblement plus souple que la nôtre.
Deux caractéristiques de la syntaxe delphinienne apparaissent néanmoins d’évidence dès la première écoute : celles de réitérance (circularité) et celle d’inclusion (enchâssement).
Lorsque le dauphin éclaire de son sonar un objet pour en saisir l’image, celle-ci est ultra-brève, juste un flash de « lumière sonore » qui dessine en un quart de seconde les contours de la cible.
Il s’agit donc de maintenir stable dans le champ mental cette réalité de type stroboscopique, ce qui n’est pas évident.
A cette fin, le cétacé va répéter et répéter encore le signal sur des modes qui varient d’un instant à l’autre, en fonction des mouvements de la cible ou de la difficulté à la circonscrire tout entière.
Le regard humain, quant à lui, « balaie » le paysage d’un mouvement presque continu. Sa vision périphérique lui assure en outre une permanence du décor dont le dauphin ne dispose pas lorsqu’il se fie à sa seule « oreille ».
Dès lors que l’homme exprime une notion (exemple : le sujet de la phrase), celle-ci persiste alors même que sa vocalisation s’est éteinte depuis longtemps. La mémoire de l’auditeur garde en mémoire chaque élément de la phrase et les relie dans son esprit pour en tirer le sens global.
Chez les dauphins, le fait d’émettre des crépitements en « loops » assortis de sons purs a pour fonction de préciser l’image stroboscopique délivrée au départ, de la rendre plus concrète par ajouts de détails partiellement identiques.
Ainsi, l’image de « ce poisson » évoqué plus haut sera répétée à de très nombreuses reprises, afin de bien définir sa taille, son
poids, son espèce et même le contenu de son estomac ou son état mental.
Des éléments anecdotiques relatifs à son comportement, aux circonstances de la rencontre seront ajoutés au fur et à mesure de la répétition du message.
La « signature sifflée », déjà longuement décrite plus haut, joue un rôle important dans le langage dauphin. Elle y revient d’une façon si fréquente que certains scientifiques ont pu croire qu’elle était la seule manifestation sifflée possible pour un dauphin. En fait, il est bien vite apparu que si les dauphins captifs utilisaient sans cesse leur signature sifflée, c’est parce qu’ils lançaient un appel au secours !
En outre, les recherches actuelles révèlent des nuances dans l’émission de ces signatures, inaccessibles aux premiers enregistrements, et montrent notamment que cette « signature » peut porter bien des informations.
III. Une liguistique universelle

A. La fonction du langage
Il est curieux que les vocalisations si complexes des cétacés et du dauphin tout particulièrement, n’ait jamais donné lieu à un raisonnement darwinien, pourtant universellement pratiqué quand il s’agit d’autres aspects du comportement animal. Cette analyse prend pour cible une fonction précise le vol, la marche debout, la chasse en groupe ou tout autre aspect du comportement et tente de comprendre pourquoi, en termes d’évolution, cette fonction a pu voir le jour.
En d’autres termes, à quoi elle sert et d’où elle vient.
Chez les humains, par exemple, la marche debout et son corollaire, la course, est le premier trait qui distingue de façon nette les hominiens naissants de tous les autres grands singes de l’époque.
Cette caractéristique a permis par la suite la survie des gènes des meilleurs coureurs, donc des meilleurs chasseurs, et a imposé au fil de la sélection naturelle une stature de plus en plus droite, une colonne vertébrale en s, un bassin aligné sur les épaules, et finalement, des mains désormais libérées aptes à toutes les technologies.
Mais le langage ? D’où provient-il et quel avantage est-il sensé offrir en termes d’évolution ?
Selon le paléontologue Richard Leakey (in « L’Origine de l’Humanité » Hachette, Paris, 2000) :
«On peut supposer que le langage ne fit pas son apparition d’un seul coup, aussi devons-nous nous demander quels bénéfices purent tirer nos ancêtres d’un langage moins développé que celui que nous connaissons.
La réponse la plus évidente est que le langage représentait un moyen de communication efficace. Cette capacité de communiquer aurait été certainement utile à nos ancêtres lorsqu’ils adoptèrent un mode subsistance fondé sur la pratique d’une chasse rudimentaire et de la cueillette, plus difficile que celui de nos grands singes.
Leur mode de vie devenant plus complexe, la coordination sociale et économique serait devenue pour eux une nécessité. Dans ces circonstances, ils auraient eu besoin d’un moyen de communication efficace. La sélection naturelle aurait donc régulièrement augmenté leur capacité de communiquer par le langage ».
Il est frappant que l’on puisse utiliser pratiquement les mêmes termes en ce qui concerne les dauphins.
Seuls de tous les mammifères, à l’exception de l’homme, ils ont développé un système phonatoire d’une formidable sophistication, les sacs nasaux logés sous l’évent, qui permet des nuances vocales bien supérieurs nombre aux cinquante phonèmes produit par notre gorge.
Par ailleurs, le développement du langage a entraîné, comme chez l’homme, un développement important du cerveau.
A cet égard, Harry Jerison accorde au langage un rôle moteur dans la croissance du cerveau et rejette l’idée selon laquelle les capacités de manipulation auraient pu être à l’origine du développement de l’encéphale – hypothèse de l’homme faiseur d’outils.
« Cette explication me semble impossible, en particulier parce qu’il est possible de fabriquer des outils avec très peu de matière cérébrale » a-t-il déclaré en 1991, « alors que la production d’un discours simple, utile, elle, exige une quantité substantielle de matière cérébrale ».
Ailleurs, le même Jerison rappelait que si le cerveau du dauphin était tout entier dévolu à l’écholocation, comme le prétendent encore quelques cétologues de l’école anthropocentriste, on se demande pourquoi les chauves-souris, qui disposent du même outillage sensoriel que le dauphin à ce niveau, ait une tête aussi petite…
L’examen neurologique nous confirme en effet que si le cerveau de certains cétacés est devenu énorme, ce n’est pas au niveau des fonctions motrices mais bien à celui des « zones silencieuses associatives», typiquement dévolues à la pensée abstraite, au sens du moi auto-réflexif et au maniement du langage.
Pourquoi ? Au fil de quelle évolution ?
Comme l’homme, le dauphin vit dans une société de chasseurs-cueilleurs, tout à la fois fondée sur l’exploitation cynégétique d’un gibier de taille moyenne (mulets, harengs, pieuvres, calmars) et sur les ressources des fonds marins (invertébrés, mollusques, petits poissons plats et autres fruits de mer cachés sous le sable.)
Leurs opérations de chasse exigent, elles aussi, une parfaite coordination à longue distance, de la même manière que nos ancêtres humains ont du peaufiner les interpellations qu’ils se lançaient d’une colline à l’autre, afin de réduire les erreurs d’interprétation.
On s’explique mieux dès lors le développement du langage chez le dauphin Tursiops ou chez l’orque : au fil des millénaires, la sélection naturelle a favorisé, chez eux aussi, la production de sons de
plus en plus complexes, tandis que se modifiait en conséquence leurs organes phonatoires.
On comprendrait mal en effet qu’une telle capacité de produire des sons aussi distincts que les clicks, braiments, «burst-pulsed sounds» et autres vocalisations delphiniennes, n’ait pas, en termes d’évolution, présenté un quelconque avantage génétique.
La complexité du monde marin, la multitude des défis qu’il impose à ceux qui vivent en son sein et la richesse de l’appareil cognitif des cétacés expliquent certainement la présence de ce véritable «langage ».
Que ce mode d’expression ne soit lié en rien aux manipulations d’outils ou à un savoir technologique nous place évidemment devant le mystère d’une pensée acoustique totalement différente, où les «mots » sont visibles et où les choses portent des «noms naturels», comme en rêvaient les philosophes du Moyen Age !
Lire à ce propos :
Cetaceans Have Complex Brains for Complex Cognition
B. L’Equivalence humaine
Il est évidemment impossible de trouver dans nos langues humaines le moindre équivalent de ces structures linguistiques. Trop de paramètres psychosensoriels, trop de données environnementales divergent dès l’abord pour rendre pertinentes de telles comparaisons.
Selon H. Jerison, par exemple, la notion même d’un « Je » central – autrement dit d’un sujet du discours – n’existe pas chez les dauphins. Ceux-ci vivent dans un monde tridimensionnel, non-soumis à la
gravité et leur concept d’individualité est sans doute altéré par le fait que les contenus mentaux de chacun sont la propriété de tous.
Jim Nollman rappelle par ailleurs, dans l’un de ses courriers daté de mars 98 que si les cachalots – dont le cerveau est cinq fois plus volumineux que le nôtre – ont bien cinq fois notre intelligence, alors il nous est tout simplement impossible de jamais les comprendre.
« Le théorème de Gödell, précise Jim Nollman, stipule qu’un traiteur de données (processor) ne peut jamais prendre en compte les capacités d’un autre traiteur de données plus important que lui « .
Autrement dit, il est hors de question pour un enfant de cinq ans d’apprécier la saveur des théories d’Einstein, hors de question pour un singe de saisir le sens d’un livre imprimé et (peut-être) impossible pour un humain adulte de suivre le fonctionnement ultrarapide de la pensée delphinienne.
Quant au contenu même de la langue delphinienne, aux valeurs qu’elle transporte, aux catégories sur lesquelles elle s’appuie, aux informations qu’elle véhicule, il est bien évident que nous ne disposons jusqu’à présent d’aucune information fiable ou qui ne soit le fruit de notre imagination.
A la manière d’une « Pierre de Rosette », les langues sifflées intermédiaires (notamment celles conçues par R.Busnel, John Lilly ou Dwight Wayne-Batteau) nous permettront sans doute un jour de
dépasser cette ignorance et de pénétrer enfin de plain-pied dans le monde mental des cétacés.
En attendant, on ne peut que s’interroger sur les messages transmis par ces dialogues entre dauphins.
Comment fait-on pour retrouver son chemin en pleine mer et sous l’eau ? Comment pose-t-on des repères dans un monde fluide ? Comment indique-t-on à quelqu’un où se trouve quelque chose, quand tout bouge sans cesse, qu’il n’y a pas de murs, d’espaces clos, et que l’on se déplace dans les trois dimensions ? Comment se souvient-on de ses voisins et amis alors qu’ils se ressemblent à l’identique comme se ressemblent deux poissons ? Ce n’est sûrement pas une tâche facile ! Et pourtant, les dauphins y parviennent.
Cela dit, leurs concepts sont-ils nécessairement incompréhensibles au regard de nos critères mentaux, du seul fait qu’ils proviennent d’une espèce aquatique dotée de nageoires plutôt que de bras ou de jambes ? Rien ne nous oblige à le croire.
On se reportera utilement à notre page intitulée « Dauphin : vie quotidienne » et l’on y verra comment l’étude objective des moeurs des cétacés mène à la conclusion que leur culture est proche de celles de nos Peuples Premiers, chasseurs-cueilleurs de la forêt pluviale ou bien aborigènes du Busch australien.
Comme eux, les dauphins vivent hors de l’histoire, dans un temps suspendu où les choses ont leur place depuis l’éternité. Comme eux, ils ne travaillent – c’est à dire « ils ne chassent » – que quelques heures par jour et passent le reste du temps à « socialiser » les uns avec les autres, à jouer, faire l’amour et tisser dans les fonds clairs des baies des chapelets de bulles artistiques.
Comme eux enfin, le plus clair de leur intelligence et de leur curiosité s’applique non pas à changer le monde environnant mais à mieux le comprendre et à bénéficier paisiblement de ses ressources.
Il est piquant à cet égard de constater qu’en 1960, un ethnologue comme Claude Levi-Strauss en était encore à plaider cette vision des choses à propos de nos « primitifs ».
La chose nous paraît aberrante puisque aujourd’hui, nul n’ignore que les peuples premiers ont développé des savoirs authentiques et puissants, simplement différents des nôtres et fondés sur d’autres paradigmes.
On sait qu’il faut autant d’intelligence, de mémoire et de sagacité à un pilote d’avion de chasse pour guider son avion qu’à un indien Yanomami pour viser et tuer un singe hurleur en pleine forêt d’un coup de sarbacane.
Que dire alors des redoutables techniques de chasse élaborées par les orques ? Des rideaux de bulles tissées par les baleines ?
Du « tir-sonar » longuement appris auprès d’un aîné par le jeune dauphin Tursiops ? Que dire enfin des techniques de navigation que dominent parfaitement les immenses
baleines bleues ?
L’intelligence verbale des cétacés n’a que faire de nos littératures, de nos gadgets technologiques aussi nuisibles qu’inutiles ou de nos réflexions narcissiques sur le rôle majeur de l’Humanité. Elle est pour eux un instrument de contrôle de la réalité et à ce titre, remplit deux fonctions propres: la survie dans le milieu naturel et la gestion des relations sociales.
Le linguiste E.Sapir a démontré comment l’environnement physique et relationnel influait puissamment sur l’élaboration des langues parlées. Il écrit notamment à propos des Indiens Païute méridionaux de l’Arizona :
« Les habitants de ce plateau désertique usent d’un vocabulaire particulièrement riche en indicateurs topographiques, dans une proportion telle que certains mots paraissent parfois trop précis que pour avoir une valeur pratique : ligne de séparation des eaux, corniche, bas- fond sablonneux, vallée semi-circulaire, vallée circulaire ou creux, petit terrain, plat entouré de crêtes montagneuses, vallée large entourée de montagnes, plain, désert, bute, plateau, gorge sans eau, gorge avec eau, torrent, sillon creusé par la pluie, flanc de montagne l’ombre, pays ondulé avec des collines basses, etc.
Il est en effet indispensable aux habitants de cette région semi-aride et inhospitalière de posséder des références topographiques très précises, pour localiser par exemple la présence d’une source-étape lors d’un voyage ».
Le vocabulaire est donc le reflet fidèle de la complexité culturelle qui l’engendre.
Mais attention : si nos langues modernes sont indubitablement riches en termes techniques les plus variées, en revanche, leur complexité est paradoxalement bien moindre que celle des peuples dits « premiers », que peu de linguistes parviennent à dominer, tant elles sont difficiles.
Lire aussi :
le monde mental des dauphins
C. Avant de parler aux extraterrestres
Lors des premières recherches entreprises par les concepteurs du Projet Seti (Carl Sagan et John Lilly, entre autres) afin d’entrer en contact avec la vie extraterrestre par le moyen d’ondes radio, le langage des dauphins a constitué une importante base de réflexion pour concevoir une pensée non-humaine.
A l’époque, l’idée prévalait qu’en étudiant attentivement ce mode de communication « exotique » et en le comparant au nôtre, serait mise à jour une « grammaire universelle », constituée de paradigmes sous-jacents communs à l’homme et au cétacé. A partir de là, des lois linguistiques infiniment plus générales pourraient être déduites et la communication avec toute autre forme d’intelligence non-humaine deviendrait nettement plus aisée. Il semble que cette idée ne soit pas abandonnée, puisqu’une expédition a été menée récemment en mer par des membres de SETI en vue d’enregistrer les chants des baleines.
On lira également l’intéressant article de John Elliott, B.Sc. (University of Leeds, UK) intitulé » Is Anybody Out There? The Detection of Intelligent and Generic Language-Like Features » et qui plaide en ce sens.
Il faut se souvenir que lorsque le Sanskrit est apparu dans les Belles Lettres occidentales, jusqu’alors confinées au Grec et au Latin, ce fut une petite révolution : le concept de langues indo-européennes en naquit puis, à sa suite, celle des familles linguistiques et enfin, celle d’une langue unique propre aux premiers Homo sapiens.
Jamais pourtant, un système syntaxique autre que celui de l’être humain n’a sérieusement été étudié, aucune étude de ce que pourrait être un » vocabulaire » dans ce contexte aussi étrange que les grands fonds marins.
Y a-t-il des mots chez les dauphins ? Des verbes, des adjectifs ? Ou bien est-ce vraiment « tout à fait autre chose » ?
Voilà bien des questions qui devraient en principe transporter d’enthousiasme tous les linguistes de la planète mais aussi les futurs astrobiologistes avides de parler avec ET.
Il n’en est rien, pourtant, sans doute du fait de la forte résistance anthropocentriste qui, comme nous le rappelions en commençant cet article, freine encore considérablement toute recherche objective sur les langages non-humains.
Livres à lire
Dolphin Cognition & Behaviour. A comparative Approach
par Ronald J. Schusterman, California State University, Hayward and University of California Lawrence Erlbaum associates, publishers New Jersey 1986. Principalement pour l’étonnant article de H.Jerison sur le langage chez le dauphin et sa possible perception du monde.
Sensory Abilities of Cetaceans: Laboratory and Field Evidence
sous la direction de Jeanette A. Thomas et Ronald Kastelein (Harderwijck Dolfinarium) dans la collection NATO ASI Series (Series A : Life sciences Vol.196)
Un livre fondamental, à lire d’urgence pour les célèbres articles de V. Markov dont on trouvera des extraits sur ce site mais aussi pour une autre étude de V. Markov et A. Zanin ( « The ability of bottlenose dolphins trusiops truncatus to report arbitrary information ») qui démontre que cette langue articulée véhicule aisément des concepts abstraits et des « représentations » d’objets de conscience non physiquement présents.
La structure récursive du langage, qui permet dans toutes les langues humaines d’inclure à gré des ensembles de mots au coeur des phrases, peut être apprise par des oiseaux chanteurs, alors que l’on croyait que cette caractéristique était le propre de l’homme. L’équipe du chercheur américain Timothy Gentner, de l’Université de Californie (San Diego), a réussi à apprendre à des sansonnets à reconnaître ce type de structure, selon une étude publiée dans la revue scientifique Nature. Ces résultats
viennent bouleverser la théorie selon laquelle cette structure grammaticale est le seul élément du langage qui soit spécifique à l’homme.
C’est la structure récursive qui permet d’enrichir sans fin les phrases, tout en conservant leur cohérence. Ainsi, explique le psychologue Gary Marcus (Université de New York) dans une analyse accompagnant l’étude, la phrase « l’amour conquiert tout » peut-elle s’incorporer à une autre pour devenir « Chris sait que l’amour conquiert tout ».
En répétant le schéma, on obtient par exemple: « Terry sait que Chris sait que l’amour conquiert tout ».
Et ainsi de suite. L’équipe de Gentner a fait écouter à ses sansonnets des séquences de chants (gazouillis et
jacassements) distribuées selon cette structure, et d’autres où les séquences étaient simplement ajoutées à la fin de la phrase musicale. Lorsque les étourneaux reconnaissaient une structure récursive, ils devaient piquer du bec sur un bouton, obtenant une récompense. Après des dizaines de milliers d’essais éducatifs, neuf des onze oiseaux étaient capables de distinguer les séquences structurées des autres. De précédents essais similaires sur des singes, menés en 2004, n’avaient abouti à aucun résultat, semblant corroborer la théorie de la spécificité humaine. L’étude publiée jeudi pose de nombreuses questions sur l’évolution du langage, note Gary Marcus. Des recherches devront être menées notamment pour savoir
si cette capacité est partagée par tous les animaux capables d’acquérir de nouvelles structures de vocalisation, à savoir les oiseaux chanteurs et, très certainement, les cétacés.